

Construction of RNA Interference Vector of Ericerus pela Chavannes) ws Gene and Preparation of dsRNA by Prokaryotic Expression
-
摘要:目的
构建白蜡虫(Ericerus pela)蜡酯合酶(wax synthase,WS)基因干扰载体并建立其体外dsRNA(double-stranded RNA,dsRNA)原核表达体系,低成本大量制备白蜡虫ws基因的dsRNA。
方法克隆白蜡虫蜡酯合酶基因ws片段,连入L4440载体,将重组质粒转入大肠杆菌HT115感受态细胞,经IPTG诱导获得与目的片段相对应的dsRNA。
结果白蜡虫ws基因RNA干扰(RNA interference,RNAi)载体成功构建,重组质粒转入HT115感受态细胞经IPTG诱导后菌体成功表达dsRNA,dsRNA的平均获得量1 705 ng·mL-1。
结论该研究通过原核表达白蜡虫ws基因的dsRNA,为后续利用RNAi实验研究白蜡虫ws基因功能及作用机理奠定基础。
Abstract:ObjectiveThis study aims at construct the interference vector of Ericerus pela wax synthase gene and prokaryotic expression system in vitro, and prepare a large number of double-stranded RNA (dsRNA) of E. pela ws gene at low cost.
MethodThe cloned E. pela ws gene fragment was inserted into L4440 vector to construct E. pela ws-L4440 gene interference vector. The recombinant plasmid was transformed into HT115 competent cell, then induced by IPTG to get the dsRNA corresponding to target fragment.
ResultThe interference vector of Ericerus pela ws gene was successfully constructed in vitro, and the dsRNA can also be expressed by HT115 competent cell with transformed recombinant plasmid induced by IPTG. The average production of dsRNA was 1 705 ng·mL-1.
ConclusionThe expression of the dsRNA of E. pela ws gene by prokaryotic expression system may lay foundation for using RNAi technology to study the function and mechanism of E. pela ws gene.
-
Keywords:
- Ericerus pela /
- WS /
- L4440 /
- prokaryotic expression /
- dsRNA
-
生物蜡酯普遍存在于自然界生物中,蜡酯对生物生命活动具有储能、保护等诸多功能,在生物生命活动中扮演的角色极为重要[1-3]。昆虫和植物体表的蜡酯层能对抗有害细菌及真菌入侵,同时还能减少体表水分蒸发、疏水御潮、防止紫外辐射、反射阳光辐射、躲避天敌袭击[4-7]。有些昆虫则特化了泌蜡的性状,比如介壳虫,这一类昆虫体表覆盖大量蜡泌物形成的介壳[8],由于蜡酯的保护,常规方法很难达到防治效果,喷洒、涂抹等机械方法和化学治理方法也较为困难不容易操作,且污染环境、耗资大、大量杀伤天敌,介壳虫已成为农林果树危害严重的害虫。如果对蜡酯进行破坏,则易达到介壳虫防治的效果。
用于昆虫实验研究的dsRNA合成方法,主要是试剂盒体外转录以及原核菌体表达获得,其中,dsRNA合成试剂盒价格较高,合成dsRNA的量较少。因此,本研究将选取前期筛选的白蜡虫ws基因片段,采用原核菌体表达合成dsRNA的方法,利用原核表达系统制备dsRNA,以期较低成本获取大量dsRNA。
不同物种的蜡酯合酶(wax synthase, WS)研究显示,在生物蜡酯合成过程中WS具有关键作用[9-12]。笔者以我国历史悠久的特产资源昆虫—白蜡虫(一种介壳虫)为研究对象,鉴定出白蜡虫WS,实验表明WS在白蜡虫泌蜡中发挥极其重要的作用[13]。近年来,昆虫中对未知基因功能及作用机理研究主要是利用dsRNA诱导的RNAi实现[14],由此本研究欲探索较低成本获取dsRNA,以期建立一种通过原核菌体表达获取大量白蜡虫ws基因dsRNA的方法,从而为介壳虫防治提供一定参考。
1. 实验材料
本实验所用白蜡虫雄虫采自中国林业科学研究院资源昆虫研究所,载体L4440及菌株HT115均为新疆大学马纪教授赠送,Escherichia coli JM109感受态细胞、Competent Cell Preparation Kit、琼脂糖凝胶电泳Marker及Loading Buffer购自大连宝生物工程有限公司;Q5® High-Fidelity DNA Polymerase、XbaⅠ及Hind Ⅲ限制性核酸内切酶购自美国NEB;M-MLV cDNA合成试剂盒、RNaseOUTTM核酸酶抑制剂、琼脂糖凝胶回收试剂盒、质粒小提试剂盒、RNase A、DNase I、Trizol均购自Thermo Fisher Scientific,InvitrogenTM(美国);焦碳酸二乙酯(Diethy pyrocarbonate, DEPC)、Luria-Bertani(LB)固体和液体培养基、氨苄青霉素(Ampicillin, AMP)、四环素(Tetracyclines, TET)、5-溴-4-氯-3-吲哚-β-D-糖苷(5-Bromo-4-chloro-3-indoxyl-α-D-galactopyranoside, X-gal)、溶菌酶、异丙基-β-D-硫代半乳糖苷(Isopropyl β-D-1-Thiogalactopyranoside, IPTG)购自生工生物工程(上海)股份有限公司;琼脂糖粉购自法国Biowest;pGEM®T-Easy Vector System、T4 DNA连接酶购自美国Promega;无水乙醇、氯仿、异丙醇、甘油购自北京国药集团化学试剂有限公司。
2. 研究方法
2.1 白蜡虫总RNA提取和引物设计
取适量白蜡虫雄虫于已灭菌离心管中,加入Trizol匀浆,提取白蜡虫雄虫总RNA。用M-MLV cDNA合成试剂盒反转录得到单链cDNA。利用软件Primer 5.0设计引物,在上游引物的5’端和下游引物3’端分别加上Hind Ⅲ、XbaⅠ酶切位点。
2.2 白蜡虫ws基因片段的克隆
以反转录获得的cDNA为模板,通过PCR扩增白蜡虫ws基因片段,利用未加酶切位点的引物组合进行扩增,反应体系分别为10 μL: 5×Q5 Reaction Buffer 2 μL、10 mmol·L-1 dNTPs 0.2 μL、Q5超保真DNA聚合酶0.1 μL、模板0.2 μL、上下游引物各0.5 μL、ddH2O补足至10 μL。扩增程序:98℃预变性30 s;98℃变性8 s,58℃退火20 s,72℃延伸15 s,30个循环;72℃延伸2 min。
37℃恒温培养12 h,挑取白色单菌落,培养后进行菌液PCR检测,同时提取重组质粒,将鉴定的阳性单克隆菌液送至昆明硕擎生物科技有限公司测序。
获得的PCR产物经1.2%的琼脂糖凝胶电泳检测鉴定,同时利用超微量分光光度计(Thermo, 美国)初步测定其浓度。检测正确的PCR产物经稀释作为模版,用带酶切位点的引物进行PCR扩增,并对目的片段进行胶回收及纯化,与质粒pGEM®T-Easy Vector连接,重组质粒命名为pGEM/EpelWS。重组质粒转化JM109感受态细胞,涂布于LB筛选平板(含有100 μg·mL-1氨苄青霉素、20 mg·mL-1 X-gal及24 mg·mL-1 IPTG)。
2.3 白蜡虫WS RNA干扰载体的构建
利用Competent Cell Preparation Kit制备试剂盒,按照说明书操作制备HT115感受态细胞。将L4440/EpelWS重组质粒转入HT115感受态细胞,涂布于含氨苄青霉素和四环素抗性的LB固体培养基。37℃过夜培养并挑取阳性单克隆,进行菌液PCR检测和双酶切检测。
测序正确的pGEM/EpelWS重组质粒和L4440载体分别提取质粒,同时利用XbaⅠ和Hind Ⅲ 37℃过夜双酶切,琼脂糖凝胶电泳检测并回收目的片段和L4440载体,二者经T4连接酶(Promega, 美国)4℃过夜连接,重组质粒转入JM109感受态细胞,涂布于含有氨苄青霉素及四环素的LB固体培养基。37℃过夜培养,挑取白色单菌落,进行菌液PCR检测,将阳性重组质粒测序验证,重组质粒命名为L4440/EpelWS。
2.4 dsRNA的表达及纯化
将上述含有L4440/EpelWS的HT115菌液于37℃条件下100 mL培养基中扩大培养OD600=0.4左右,加入IPTG使其终浓度为0.7 mmol·L-1,继续培养过夜诱导表达dsRNA。将菌液离心收集菌体,用浓度为500 mg·mL-1的溶菌酶常温裂解菌体5 min,Trizol法提取dsRNA,用灭菌的DEPC处理水溶解dsRNA,并从溶解液中取出少量,利用超微量分光光度计测定其浓度,同时进行电泳检测。将获得的dsRNA进行DNase I和RNase A消化处理,除去其中的DNA和单链RNA,以获得较纯的dsRNA。
3. 结果与分析
3.1 目的片段扩增及阳性单克隆筛选
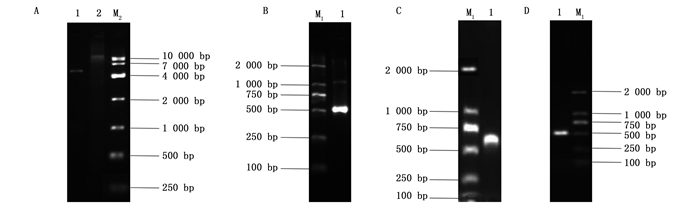
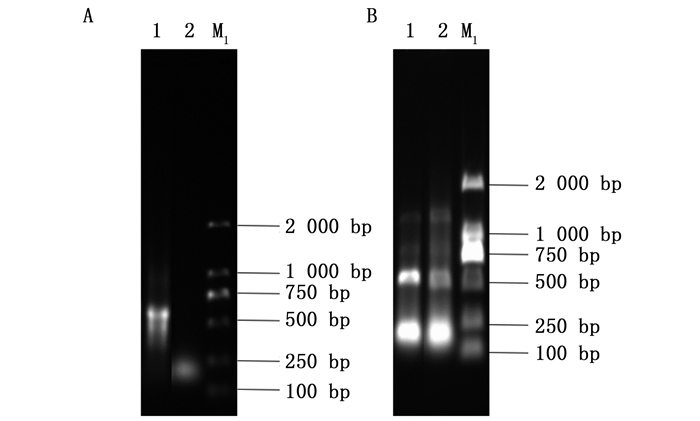
以反转录获得的cDNA为模板,通过PCR扩增白蜡虫ws基因片段约500 bp,扩增的目的片段与预期片段基本相符(图 1 A)。以此PCR扩增产物为模版,采用带XbaⅠ和Hind Ⅲ酶切位点的引物进行扩增,对所挑取的阳性重组单克隆pGEM/EpelWS菌液检测的结果也与预期片段基本相符(图 1 B),其测序结果正确。
3.2 重组载体的构建
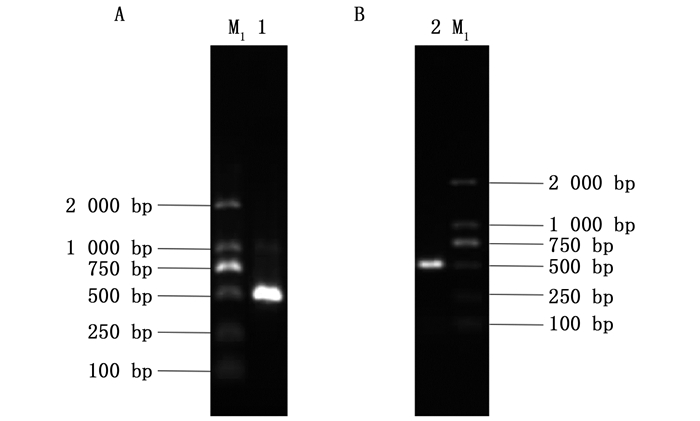
利用酶XbaⅠ和Hind Ⅲ同时对L4440质粒和pGEM/EpelWS重组质粒进行双酶切,L4440质粒双酶切的结果如图 2 A:第1个泳道是XbaⅠ和Hind Ⅲ双酶切之后的电泳结果,第2个泳道是L4440质粒电泳检测结果。pGEM/EpelWS重组质粒双酶切的结果如图 2 B,即pGEM/EpelWS重组质粒经Xba Ⅰ和Hind Ⅲ双酶切之后的电泳结果。二者经T4连接酶连接、转化,以白色单克隆的菌液为模版,特异性引物PCR扩增验证结果如图 2 C,检测结果与预期片段大小基本相符,菌液测序结果也正确。
上述测序正确的L4440/EpelWS菌液,对其提取质粒并转入HT115感受态细胞,挑取单克隆摇菌,菌液利用特异性引物检测,结果见图 2 D,也与预期片段基本相符。
3.3 原核诱导表达白蜡虫ws基因的dsRNA
![]() 图 3
图 3细菌中诱导表达dsRNA ws的鉴定
Figure 3.Identification of dsRNA of ws gene produced in bacteria
经IPTG诱导后表达的dsRNA电泳图(图 3)表明:转化HT115感受态细胞并经IPTG诱导后的L4440/EpelWS菌液成功表达了相应的dsRNA,提取dsRNA的平均浓度为1 705 ng·mL-1,而未经IPTG诱导的L4440/EpelWS菌液未表达出相应dsRNA;没有转化HT115感受态细胞的L4440/EpelWS菌液,在IPTG诱导诱导前后均未表达出dsRNA。对已经诱导表达的dsRNA进行DNase I和RNase A消化处理的电泳结果如图 3 B所示。
4. 讨论
目前,昆虫RNAi普遍利用注射法和饲喂法[15],在昆虫中利用RNAi沉默特定基因的方法已经成熟,如通过显微注射胚胎和注射若虫或成虫血腔导致基因沉默[16-18],但是注射法对昆虫有一定损伤甚至致死现象,如对黄粉虫(Tenebrio molitor)和Dendroides canadensis注射dsRNA后,注射dsRNA引起的昆虫死亡率在一定时间内呈上升趋势,同时抗冻蛋白(Antifreeze protein, AFP)基因afp表达量也有明显增加[19-20];饲喂法最先是对Rhodnius prolixus和Epiphyas postvittana进行,其结果发现RNAi效果明显可有效降低目的基因转录[21],Bilgi等[22]对蚜虫探索有效的饲喂dsRNA也明显达到蚜虫RNAi效果;此外,针对不同形态类型昆虫体表涂抹dsRNA也不失为较好的昆虫RNAi方法[23],但是这种方法需要较多的dsRNA。由于饲喂法的RNAi效果实现需用大量的dsRNA,因此,如何获取大量dsRNA是降低成本广泛应用该方法的必要条件。
由于原核菌体表达获得dsRNA成本低、获取量较高等优点,现已用于鞘翅目、直翅目等昆虫及多种生物的基因功能研究[24]。本研究采用原核菌体表达获得白蜡虫WS对应dsRNA,与试剂盒合成dsRNA相比,其优点在于仅需少量菌液,即可在短时间内诱导获取大量目的基因的dsRNA,同时其成本也大大降低。因L4440载体含有双向T7启动子和lac乳糖操纵子,大肠杆菌HT115为RNase Ⅲ缺陷型菌株,所以菌液诱导表达目的基因dsRNA不会被RNase Ⅲ酶切降解[25],故L4440用于多种生物体外目的基因载体构建获取dsRNA,如黄粉虫抗冻蛋白基因afp原核表达dsRNA重组载体构建,获得黄粉幼虫抗冻蛋白基因的dsRNA,纯化后成功对黄粉幼虫进行RNAi[20];甘蔗二点螟(Chilo infuscatellus Snellen)蜕皮调节转录因子CiHR3的RNAi载体构建,利用获得的dsRNA饲喂甘蔗二点螟幼虫,可有效抑制CiHR3的正常表达[26];中华绒螯蟹(Eriocheir sinensis)EsSox21b-like基因利用原核表达制备dsRNA[27]。
5. 结论
本研究利用原核表达系统通过细菌大量制备白蜡虫ws基因的dsRNA,dsRNA的平均获得量1 705 ng·mL-1,电泳检测显示拖尾现象不明显,dsRNA的质量较好,仅需50 mL的菌液就可获得85.25 μg·次-1,与试剂盒MEGAscriptTM RNAi Kit with Manual的50~100 μg·次-1获取量相当,可节省大量资金。后续将对研究对象白蜡虫进行RNAi操作,对白蜡虫WS功能展开深入研究,也为其他昆虫dsRNA合成提供参考和理论依据。
-
![]()
图 1
目的片段克隆结果
A:为不含酶切位点的pGEM/EpelWS电泳检测结果;B:pGEM/EpelWS菌液PCR检测;M1: DL2000 DNA Marker Figure 1.
The cloning results of target fragment
A. 1: the electrophoresis detection results of pGEM/EpelWS without restriction site; B. 2: PCR detection of pGEM/EpelWS bacterium liquid; M1: DL2000 DNA Marker ![]()
图 2
载体构建
A: 1为载体L4440质粒经Xba Ⅰ和Hind Ⅲ双酶切之后的结果,2为L4440质粒电泳检测;B: 1为带Xba Ⅰ和Hind Ⅲ酶切位点的pGEM/EpelWS重组质粒经Xba Ⅰ和Hind Ⅲ双酶切之后的结果;C: 1为将目的片段连入L4440载体转化JM109感受态细胞后,菌液PCR检测结果;D: 1为L4440/EpelWS质粒转入HT115感受态细胞,经特异性引物PCR扩增验证转化的菌液检测结果; M1: DL2000 DNA Marker; M2: DL10000 DNA Marker Figure 2.
The construction of the interference vector
A. 1: the result of L4440 plasmid vector after double enzyme digestion with Xba Ⅰ and Hind Ⅲ; A. 2: the electrophoresis detection of L4440 plasmid; B. 1: The result of pGEM/EpelWS recombinant plasmit with Xba Ⅰ and Hind Ⅲ restriction site after double enzyme digestion with Xba Ⅰ and Hind Ⅲ; C. 1: PCR detection results of bacterium liquid after target fragment was inserted into L4440 and transformed the JM109 competent cell; D. 1: detection results of bacterium liquid transformed by L4440/EpelWS plasmid transferred into HT115 competent cell after specific primers PCR amplification and verification; M1: DL2000 DNA Marker; M2: DL10000 DNA Marker ![]()
图 3
细菌中诱导表达dsRNA ws的鉴定
A: 1即L4440/EpelWS菌液经IPTG诱导后提取的dsRNA,2即L4440/EpelWS菌液未经IPTG诱导提取dsRNA结果;B: 1是经DNase I消化处理之后的电泳结果,2是经RNase A消化处理之后的电泳结果; M1: DL2000 DNA Marker Figure 3.
Identification of dsRNA of ws gene produced in bacteria
A. 1: the dsRNA extracted from L4440/EpelWS bacterium liquid induced by IPTG; A. 2: the dsRNA extracted from L4440/EpelWS bacterium liquid without induction by IPTG; B. 1: extraction product of dsRNA treated by DNase I; B. 2: extraction product of dsRNA treated by RNase A; M1: DL2000 DNA Marker -
[10] 刘博文, 王雪庆, 孙涛, 等.白蜡虫蜡酯合酶基因cDNA全长克隆及原核表达[J].林业科学研究, 2016, 29(4):610-614.
DOI: 10.3969/j.issn.1001-1498.2016.04.022[14] Zhu F, Xu J, Palli R, et al. Ingested RNA interference for managing the populations of the Colorado potato beetle, Leptinotarsa decemlineata[J]. Pest Management Science, 2011, 67(2):175-182.
DOI: 10.1002/ps.v67.2[25] Newmark P A, Reddien P W, Cebrià F, et al. Ingestion of bacterially expressed double-stranded RNA inhibits gene expression in planarians[J]. Proceedings of the National Academy of Sciences of the United States of America, 2003, 100(Suppl):11861-11865.
http://d.old.wanfangdata.com.cn/OAPaper/oai_pubmedcentral.nih.gov_304099[7] 杨璞, 徐冬丽, 陈晓鸣, 等.蜡酯合成途径及关键酶的研究进展[J].中国细胞生物学学报, 2012, 34(7):695-703.
http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=xbswxzz201207010[11] Cheng J B, Russell D W. Mammalian wax biosynthesis. Ⅰ. Identification of two fatty acyl-coenzyme a reductases with different substrate specificities and tissue distributions[J]. Journal of Biological Chemistry, 2004, 279(36):37789-37797.
DOI: 10.1074/jbc.M406225200[19] Wang L. Identification and characterization of protein enhancers of antifreeze proteins from overwintering beetle larvae Dendroides canadensis[J]. Dissertations & Theses-Gradworks, 2005, 8(2):1120-1120.
[5] Reiser S, Somerville C. Isolation of mutants of Acinetobacter calcoaceticus deficient in wax ester synthesis and complementation of one mutation with a gene encoding a fatty acyl coenzyme A reductase[J]. Journal of Bacteriology, 1997, 179(9):2969-2975.
DOI: 10.1128/jb.179.9.2969-2975.1997[6] 谢映平, 薛皎亮, 郑乐怡.云南双蜡蚧蜡泌物的超微形态与化学成分[J].昆虫学报, 2004, 47(3):320-328.
DOI: 10.3321/j.issn:0454-6296.2004.03.008[26] 张雨良, 黄启星, 张树珍, 等.细菌介导RNAi研究甘蔗二点螟蜕皮调节转录因子CiHR3基因功能[J].应用昆虫学报, 2013, 50(05):1301-1310.
http://d.old.wanfangdata.com.cn/Conference/7933033[8] 谢映平, 薛皎亮, 张艳峰, 等.蚧虫蜡泌物的化学研究进展[J].应用昆虫学报, 2004, 41(6):512-518.
DOI: 10.3969/j.issn.0452-8255.2004.06.003[21] Araujo R N, Santos A, Pinto F S, et al. RNA interference of the salivary gland nitrophorin 2 in the triatomine bug rhodnius prolixus (Hemiptera:Reduviidae) by dsRNA ingestion or injection[J]. Insect Biochemistry & Molecular Biology, 2006, 36(9):683-693.
http://d.old.wanfangdata.com.cn/Periodical/syktykf201704014[9] 亓倩, 于淑惠, 孙涛, 等.白蜡虫蜡酯合酶在昆虫细胞Sf9中的表达[J].林业科学研究, 2016, 29(2):191-195.
DOI: 10.3969/j.issn.1001-1498.2016.02.006[24] Ratzka C, Gross R, Feldhaar H. Systemic gene knockdown in Camponotus floridanus, workers by feeding of dsRNA[J]. Insectes Sociaux, 2013, 60(4):475-484.
DOI: 10.1007/s00040-013-0314-6[22] Bilgi V, Fosunyarko J, Jones M G. Using vital dyes to trace uptake of dsRNA by green peach aphid allows effective assessment of target gene knockdown[J]. International Journal of Molecular Sciences, 2017, 18(1):80.
DOI: 10.3390/ijms18010080[23] Wang Y, Zhang H, Li H, et al. Second-generation sequencing supply an effective way to screen RNAi targets in large scale for potential application in pest insect control[J]. Plos One, 2011, 6(4):e18644.
DOI: 10.1371/journal.pone.0018644[13] Yang P, Zhu J Y, Gong Z J, et al. Transcriptome analysis of the Chinese white wax scale Ericerus pela with focus on genes involved in wax biosynthesis[J]. Plos One, 2012, 7(4):e35719.
DOI: 10.1371/journal.pone.0035719[1] Cheng J B, Russell D W. Mammalian wax biosynthesis. Ⅱ. Expression cloning of wax synthase cDNAs encoding a member of the acyltransferase enzyme family[J]. Journal of Biological Chemistry, 2004, 279(36):37798-37807.
DOI: 10.1074/jbc.M406226200[17] Bucher, G, Scholten J, Klingler M, et al. Parental RNAi in Tribolium (Coleoptera)[J]. Current Biology Cb, 2002, 12(3):85-86.
DOI: 10.1016/S0960-9822(02)00666-8[2] Stöveken T, Kalscheuer R, Malkus U, et al. The wax ester synthase/acyl coenzyme A:diacylglycerol acyltransferase from Acinetobacter sp. strain ADP1:characterization of a novel type of acyltransferase[J]. Journal of Bacteriology, 2005, 187(4):1369-1376.
DOI: 10.1128/JB.187.4.1369-1376.2005[3] 孙涛, 王雪庆, 赵遵岭, 等.越冬白蜡虫微生物多样性分析[J].林业科学研究, 2017, 30(6):1009-1014.
http://www.lykxyj.com/ch/reader/view_abstract.aspx?flag=1&file_no=20170618&journal_id=lykxyj[16] Kennerdell J R, Yamaguchi S, Carthew R W. RNAi is activated during Drosophila oocyte maturation in a manner dependent on aubergine and spindle-E[J]. Genes & Development, 2002, 16(15):1884-1889.
DOI: 10.1101-gad.990802/[4] 刘魏魏, 杨璞, 陈晓鸣.白蜡虫热激蛋白基因在低温胁迫下的表达分析[J].林业科学研究, 2013, 26(6):681-685.
http://www.lykxyj.com/ch/reader/view_abstract.aspx?flag=1&file_no=20130602&journal_id=lykxyj[15] Terenius O, Papanicolaou A, Garbutt J S, et al. RNA interference in Lepidoptera:an overview of successful and unsuccessful studies and implications for experimental design[J]. Journal of Insect Physiology, 2011, 57(2):231-245.
DOI: 10.1016/j.jinsphys.2010.11.006[27] 刘志强, 陈洁, 邱高峰.中华绒螯蟹EsSox21b-like基因干扰载体的构建及原核表达制备dsRNA[J].生物技术通报, 2014(6):134-138.
http://d.old.wanfangdata.com.cn/Periodical/swjstb201406022[18] Tomoyasu Y, Denell R E. Larval RNAi in Tribolium (Coleoptera) for analyzing adult development[J]. Development Genes & Evolution, 2004, 214(11):575-578.
http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=dbe692bce6c209d4092b8ffa0d798a62[12] Teerawanichpan P, Qiu X. Fatty acyl-CoA reductase and wax synthase from Euglena gracilis in the biosynthesis of medium-chain wax esters[J]. Lipids, 2010, 45(3):263-273.
DOI: 10.1007/s11745-010-3395-2[20] 石萌, 刘小宁, 马纪, 等.利用细菌表达dsRNA介导黄粉虫抗冻蛋白基因的RNA干扰[J].生物技术通报, 2014, 46(8):113-119.
http://d.old.wanfangdata.com.cn/Periodical/swjstb201408019
 下载:
下载:
计量
- 文章访问数: 5010
- HTML全文浏览量: 2026
- PDF下载量: 450
 下载:
下载:












