

Cloning and Expression Pattern of Sensory Neuron Membrane Protein Genes of Tropidothorax elegans
-
摘要:目的
感觉神经元膜蛋白(SNMP)是至关重要的次要嗅觉蛋白,能够作为信息素识别的标记,而红脊长蝽(Tropidothorax elegans)作为一个重要的农林害虫,这方面的研究还很少。
方法利用PCR方法对红脊长蝽SNMP基因进行克隆,并通过荧光定量PCR对该基因的表达情况进行分析。
结果首次克隆和鉴定了红脊长蝽的2个SNMPs基因,并命名为TeleSNMP1和TeleSNMP2。序列分析表明,TeleSNMP1编码区开放阅读框长1 497 bp,编码498 aa;而TeleSNMP2编码区开放阅读框长1 686 bp,编码561 aa,这2个基因的等电点分别为8.26和7.05。同源性分析发现,同目昆虫同类SNMP序列一致性较高,不同目昆虫不同类SNMP序列一致性较低。TeleSNMP1和TeleSNMP2之间的序列一致性极低。进化树结果也表现同目昆虫同类SNMP基因进化关系最近。定量PCR结果显示,TeleSNMP1主要在雌雄触角中表达,而TeleSNMP2在非触角组织中也有表达。
结论该结果为今后对红脊长蝽SNMPs基因功能的研究提供了有用的参考。
Abstract:ObjectiveTo study the sensory neuron membrane proteins (SNMPs) in Tropidothorax elegans.
MethodThe SNMP genes were cloned by PCR, and the expression of these genes were analyzed by fluorescence quantitative PCR.
ResultTwo SNMPs genes of T. elegans, named as TeleSNMP1 and TeleSNMP2, were cloned and identified for the first time by RT-PCR. Sequence analysis showed that the TeleSNMP1 encoding area was 1 497 bp in length, encoding 498 aa. The TeleSNMP2 encoding area was 1 686 bp in length, encoding 561 aa. The isoelectric points of these two genes were 8.26 and 7.05, respectively. Homology analysis showed that the consistency of SNMPs genes in insects of the same order was high, while the consistency of different order was low, and the sequence consistency between TeleSNMP1 and TeleSNMP2 was extremely low. The evolutionary tree results also showed that the evolutionary relationship of SNMPs genes in insects of the same order was rather close. Quantitative real-time PCR showed that TeleSNMP1 was antennae-biased, whereas TeleSNMP2 was expressed in female and male antennae but was also expressed in nonantennal tissues.
ConclusionThe results provide a useful reference for future studies on the function of the T. elegans SNMPs gene.
-
昆虫SNMPs也是一种膜蛋白,与脊椎动物CD36家族为同源基因,具有2个跨膜区域,其功能主要是识别和转运亲脂性气味分子如脂肪酸和脂类化合物等[9-14]。昆虫第一个SNMP基因在多音天蚕(Antheraea polyphemus Cramer)中被鉴定,并命名为ApolSNMP1[9]。随后在烟草天蛾(Manduca sexta L.)中发现SNMP的第二个亚类型,即命名为MsexSNMP2[10, 15]。紧接着SNMP的同源基因在鳞翅目Lepidoptera[10, 15-16]、双翅目Diptera[17]、鞘翅目Coleoptera[18]、直翅目Orthoptera[13]和膜翅目Hymenoptera[19]等昆虫中都有发现。一直以来被认为SNMP基因家族就2个成员,即SNMP1和SNMP2。最近昆虫SNMP家族的第三个成员SNMP3在鳞翅目中被鉴定[20-21],但认为该基因的主要功能与昆虫的免疫反应有关,这还需要进一步的去证明。
红脊长蝽(Tropidothorax elegans Distant)属半翅目(Hemiptera)长蝽科(Lygaeidae),主要为害刺槐(Robinia pseudoacacia L.)、辣椒(Capsicum annuum L.)、葫芦(Lagenaria siceraria Molina)、油菜(Brassica napus L.)、大白菜(Brassica pekinensis Lour.)和小麦(Triticum aestivum L.)等多种植物,食性较杂。关于红脊长蝽的嗅觉基因研究较少,本研究通过前期红脊长蝽触角转录组测序结果[22],鉴定了红脊长蝽的2个SNMP基因,即TeleSNMP1和TeleSNMP2,并通过荧光定量PCR技术对红脊长蝽TeleSNMP1和TeleSNMP2在不同组织中的表达情况进行分析,为进一步探索红脊长蝽SNMPs的化学通讯功能奠定基础。
昆虫的化学感受系统在其生存和繁殖过程中起着极其重要的作用[1-3]。在过去10多年的时间里,研究者对昆虫触角嗅觉信号传导的分子机制研究有了突出的进步。在昆虫嗅觉识别时,气味分子从触角感器孔渗入,然后被气味结合蛋白(odorant binding proteins,OBPs)或化学感受蛋白(chemosensory proteins,CSPs)识别和转移,最后激活位于嗅觉感觉神经元(olfactory sensory neurons,OSNs)树突膜上的嗅觉受体(odorant receptors,ORs)或离子型受体(ionotropic receptors,IRs),产生电位,指导昆虫做出相应的行为反应[4-8]。此外,还有一些蛋白如感觉神经元膜蛋白(sensory neuron membrane proteins,SNMPs)在昆虫气味识别过程中也扮演着至关重要的作用[5]。
1. 材料与方法
1.1 试虫的准备
红脊长蝽来自河南科技大学林学院昆虫实验室,该群体为自2014年7月在洛阳周边(112˚26′ E,34˚43′ N)蔬菜地采集的成虫,然后在温室内继代饲养至今。温室条件为:温度25 ± 2℃,相对湿度60% ± 5%,光周期14L∶10D。
1.2 总RNA的提取与第一链cDNA的合成
选取红脊长蝽羽化后第3 d的雌雄成虫,收集触角各100头、头部各30头、胸部各20头、腹部各5头、足各50头、翅各50头,每个样品收集材料重复3次。将红脊长蝽各部分组织解剖后立即放入浸在液氮中的1.5 mL离心管内,然后保存于−80℃中。
总RNA的提取采用RNAiso Plus Kit(TaKaRa,北京)试剂盒进行,并使用RNase-free DNase I(TaKaRa,北京)对提取的RNA进行除DNA处理。采用1%的琼脂糖凝胶电泳和NanoDrop 2000c分光光度计(Thermo Scientific)进行质量检测。采用PrimeScript™ 1st Strand cDNA Synthesis Kit(TaKaRa,北京)试剂盒对RNA进行反转录。
1.3 红脊长蝽SNMPs基因的克隆
从本实验室前期对红脊长蝽触角转录组测序注释结果中[21]搜素到2个SNMP基因。根据该序列设计特异性引物,TeleSNMP1-F:5′-ATGGCTGCACCACTGAGG-3′,TeleSNMP1-R:5′- CTAGTACTTTGCCGGGGGTG-3′;TeleSNMP2-F:5′-ATGACGAAGGTGCTGTTCCC-3′,TeleSNMP2-R:5′-TTAGCTTGTGAGAGTCCTTTTGA-3′,进行PCR扩增。PCR反应体系为20 μL:雌蛾触角cDNA模板1 μL,上下游引物各1.5 μL(10 μmol·L−1),dNTPs混合液1.6 μL(2.5 μmol·L−1),Ex Taq DNA聚合酶0.2 μL(TaKaRa,大连),10 × Ex Taq buffer 2 μL,ddH2O为12.2 μL。PCR反应条件为:94℃ 5 min;94℃ 30 s,58℃ 30 s,72℃ 30 s,35个循环;最后72℃ 10 min。胶回收目的片段,将目的片段克隆到pMDTM19-T载体上,转化DH5α感受态细胞,挑取阳性克隆培养过夜,送去测序。
1.4 红脊长蝽SNMPs基因的序列分析
核酸序列采用在线工具(http://www.bio-soft.net/sms/)翻译;采用(http://www.cbs.dtu.dk/services/TMHMM)在线工具进行跨膜区域预测;利用在线工具(https://web.expasy.org/protparam/)对蛋白序列特性进行分析;蛋白亲疏水性采用在线工具(https://web.expasy.org/protscale/)分析;序列比对采用线下DNAMAN进行多重序列比较;进化树采用线下MEGA 6.0构建。
1.5 实时定量PCR
荧光定量PCR被执行在ABI 7500 PCR仪(ABI,Carlsbad,CA,USA)上进行,每个反应体积20 μL,包括10 μL的2 × SYBR Green PCR Master Mix(TaKaRa,大连)、0.8 μL的正反向引物(10 μmol·L−1)、2 μL的cDNA模板(200 ng)和6.4 μL的DEPC水。PCR循环遵循95℃ 30 s,然后95℃ 5 s,53℃ 31 s,共循环40个周期。试验重复3次。
通过荧光定量PCR检测红脊长蝽SNMPs基因在不同组织中的表达情况。根据红脊长蝽SNMPs基因的开放阅读框和荧光定量引物设计原则设计特异性引物,TeleSNMP1-F:TCACCATCCCTCATCCA,TeleSNMP1-R:TCTTCGCCGTCTTTCAT,TeleSNMP2-F:TGTGGGAACGGAACTCT,TeleSNMP2-R:GCACCTTGGCACTTTG。内参基因为红脊长蝽Actin基因(基因登陆号为MG322127),引物为:TeleActin-F:CAAGGACGAAACAATCA;TeleActin-R:GAGAATACACTCCCAGAAC。
1.6 数据分析
红脊长蝽SNMPs基因在不同组织中的相对表达量采用公式2−∆∆CT[23]计算。利用SPSS 17.0软件中的ANOVA方法对TeleSNMPs在红脊长蝽不同组织中的表达量进行显著性差异比较(新复极差法检验,P ≤ 0.05)。
2. 结果与分析
2.1 红脊长蝽SNMPs基因克隆及序列分析
![]() 图 1
图 1红脊长蝽TeleSNMP1和TeleSNMP2与其他昆虫SNMPs的序列比对
Figure 1.Alignment of Tropidothorax elegans SNMP1 and SNMP2 with SNMPs from other insects
这2个SNMPs基因都具有全长的开放阅读框,TeleSNMP1长度为1 497 bp,编码498个氨基酸(图1);而TeleSNMP2长度为1 686 bp,编码561个氨基酸。这2个基因都是酸性,TeleSNMP1的等电点是8.26,蛋白分子量是55.59 kD;TeleSNMP2的等电点是7.05,蛋白分子量是63.37 kD。
将测序结果在NCBI上进行同源性搜索,结果表明所测序列与多种昆虫的SNMP基因序列高度同源,证明这2个基因就是红脊长蝽的SNMP基因,并分别命名为TeleSNMP1和TeleSNMP2,在NCBI基因登陆号分别为MW442946和MW442947。
TeleSNMP1和TeleSNMP2在氨基酸序列的N-端和C-端各有一个跨膜区域,其中TeleSNMP2在氨基酸第158和180之间还有一个跨膜区域。在膜外区域6个保守的半胱氨酸残基位点被预测(图1),这6个保守的半胱氨酸位点与CD36基因家族相似。
2.2 红脊长蝽SNMPs与其他昆虫SNMP基因的同源性及进化树分析
将红脊长蝽TeleSNMP基因与已报道的其他昆虫SNMP基因的氨基酸序列在NCBI中的Blastp在线搜索工具进行同源性分析,发现TeleSNMP基因与不同种类昆虫SNMP基因一致性差别较大。与同目昆虫SNMP基因一致性较高,如TeleSNMP1与茶翅蝽 (Halyomorpha halys Stal) HhalSNMP1序列一致性在81.33%;与苜蓿盲蝽 (Adelphocoris lineolatus Goeze) AlinSNMP1序列一致性在68.54%;与不同目昆虫SNMP基因一致性较低,如TeleSNMP1与斑痣悬茧蜂 (Meteorus pulchricornis Wesmael) MpulSNMP1序列一致性在43.44%。但TeleSNMP2与所有昆虫SNMP基因一致性都不高,如与茶翅蝽HhalSNMP2序列一致性在48.68%;与苜蓿盲蝽AlinSNMP2序列一致性在41.18%;与德国小蠊 (Blattella germanica Linnaeus) BgerSNMP1序列一致性在46.22%。TeleSNMP2与玉带凤蝶 (Papilio polytes L.) PpolSNMP2序列一致性在30.94%;与亚洲小车蝗 (Oedaleus asiaticus Bei-Bienko) OasiSNMP2序列一致性在34.76%。红脊长蝽TeleSNMP1和TeleSNMP2之间序列一致性仅有29.66%。
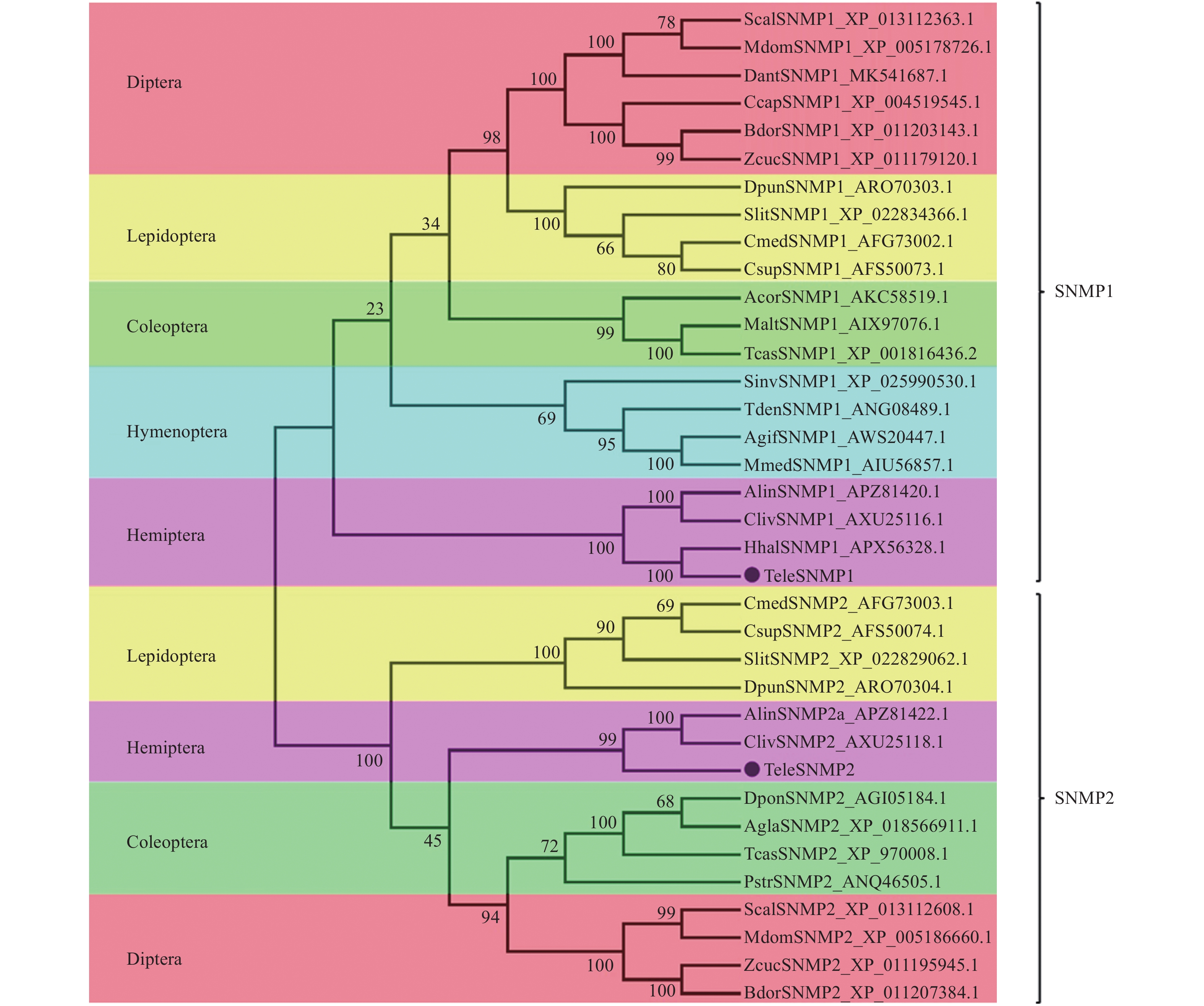
使用MEGA6.0对9个目的36种昆虫SNMP同源基因进行进化树比较。SNMPs基因被分成2个亚组,即SNMP1和SNMP2(图2),每亚组中同目昆虫SNMP基因同源性最高。红脊长蝽的TeleSNMP1和TeleSNMP2基因分别被集聚到这2个亚组,并且与同为半翅目昆虫的苜蓿盲蝽、茶翅蝽和黑肩绿盲蝽SNMP基因进化关系最近,与其他目昆虫SNMP关系较远。
![]() 图 2
图 2红脊长蝽SNMPs与其他昆虫SNMPs氨基酸序列的系统进化树
Figure 2.Phylogenetic tree of the SNMPs of Tropidothorax elegans and other insects based on amino acid sequences by using neighbor-joining method
2.3 红脊长蝽SNMPs基因表达谱分析
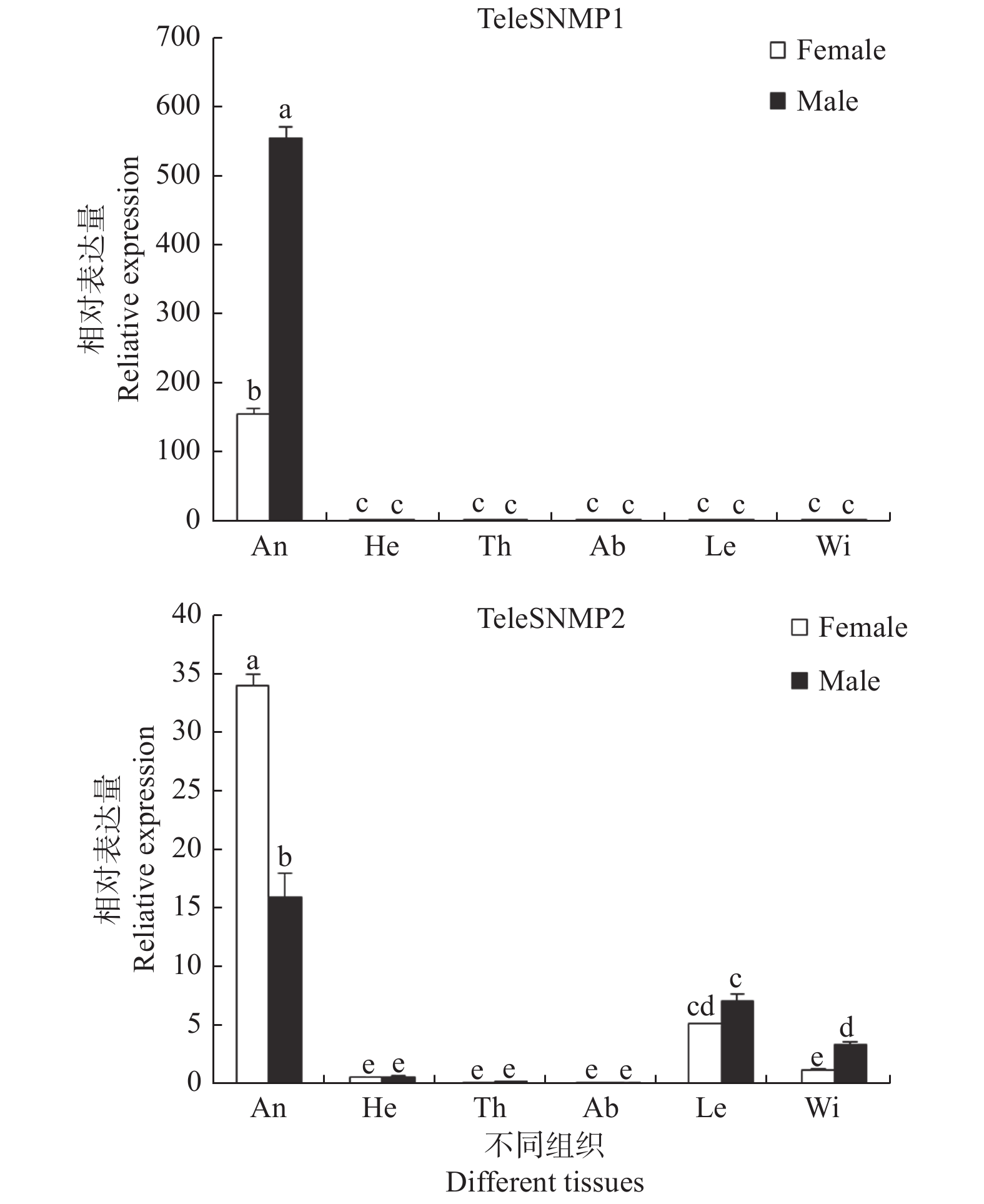
使用荧光定量PCR对红脊长蝽TeleSNMP1和TeleSNMP2在不同组织中的表达情况进行分析,结果发现:TeleSNMP1和TeleSNMP2在雌雄蛾触角中高度表达,TeleSNMP1在雄蛾触角中的表达量明显高于雌蛾,TeleSNMP2的表达情况刚好相反,即TeleSNMP2在雌蛾触角中的表达量明显高于雄蛾。除触角外,TeleSNMP1几乎不在其他组织中表达或表达量甚微。TeleSNMP2除在触角中表达量丰富外,在足和翅中也有少量表达(图3)。
![]() 图 3
图 3红脊长蝽SNMPs在不同组织中的相对表达量
Figure 3.Expression level of SNMPs in different tissues of Tropidothorax elegans
3. 讨论
红脊长蝽TeleSNMP基因同源性搜索发现TeleSNMP基因与不同种类昆虫SNMP基因一致性差别较大。与同目昆虫SNMP基因一致性较高,与不同目昆虫SNMP基因一致性较低。红脊长蝽TeleSNMP1和TeleSNMP2之间分歧也比较大。进化树结果也显示,红脊长蝽TeleSNMP1和TeleSNMP2分别被集聚到SNMP1和SNMP2两个亚组,在同一组内同目昆虫的SNMP基因进化关系最近,与其他目昆虫SNMP关系较远。此结果与大多数昆虫SNMP基因特性相同[26,29-30]。在鳞翅目中曾经鉴定出SNMP3亚家族基因[20],而在红脊长蝽触角转录组数据中没有发现该基因,可能是这个基因在幼虫肠中高度表达的原因。
组织特异性表达可以为功能预测提供可靠性的参考。我们研究发现红脊长蝽SNMP1主要表达在雌雄蛾的触角中,此结果与多音蚕蛾[9]、脐橙螟(Amyelois transitella Walker)[31]、甜菜夜蛾(Spodoptera exigua Htibner)[32]和中红侧沟茧蜂(Microplitis mediator Haliday)[33]等多种昆虫SNMP1的表达模式相同。Benton等[34]在果腹黑蝇(Drosophila melanogaster wDm)中发现,DmelSNMP1能够识别集合信息素cVA,激活受体HR13。Pregitzer等[28]在烟芽夜蛾(Heliothis virescens Fabricius)中也发现,HvirSNMP1能明显增强HR13对信息素Z11-16:Ald的结合力。以上研究都暗示昆虫SNMP1的功能是调节信息素的识别和通过SNMP-OR互作转运信息素。相对于TeleSNMP1,TeleSNMP2的表达相对广泛,除嗅觉感器触角外,在非嗅觉感器足中也有少量表达,此结果与小菜蛾(Plutella xylostella L.)[35]、二化螟(Chilo suppressalis Walker)[36]、稻纵卷叶螟(Cnaphalocrocis medinalis G.)[37]、甜菜夜蛾[31]和中红侧沟茧蜂[32]一致。昆虫身体上分布有大量的味觉感器[38],而CD36蛋白的主要功能是转运脂类化合物,因此可推测TeleSNMP2和GRs在这些组织中共同表达,识别脂类分子完成味觉过程。
本研究根据前期红脊长蝽触角转录组的数据,通过BLASTX在线搜索和同源性比较鉴定出2个SNMPs基因,即TeleSNMP1和TeleSNMP2。这2个基因与其他昆虫SNMPs具有类似的特征,如在氨基酸序列N端和C端附近有2个保守的跨膜区域,并且由6个保守的半胱氨酸残基形成二硫键组成一个大的胞外环,此结构也与CD36基因家族极其相似[24-26]。根据对这2个跨膜蛋白结构投影预测,这部分的功能是转运和结合脂类分子[27-28],我们可推测红脊长蝽SNMPs的2个跨膜区具有同样的功能。但红脊长蝽TeleSNMP2在氨基酸序列第158和180之间多了1个跨膜区域,目前关于SNMPs基因具有3个跨膜区域的报道还没有,可能是鉴定的SNMPs数量还不够庞大,也可能长期进化形成的。
4. 结论
本研究首次克隆和鉴定了红脊长蝽的2个SNMPs基因,即TeleSNMP1和TeleSNMP2。同目昆虫同类SNMP同源序列一致性较高,反之,不同目昆虫不同类SNMP同源序列一致性较低。同样,TeleSNMP1和TeleSNMP2之间的序列一致性也极低。TeleSNMP1主要在雌雄触角中特异性表达,而TeleSNMP2除触角外,在非触角组织中也有表达。
-
![]()
图 1
红脊长蝽TeleSNMP1和TeleSNMP2与其他昆虫SNMPs的序列比对
注:方块内为跨膜区域;6个保守的半胱氨酸位点用小星星表示;缩写的种名分别是:Tele,红脊长蝽;Aips,小地老虎;Alin,苜蓿盲蝽;Slit,斜纹夜蛾;Btab,烟粉虱. Figure 1.
Alignment of Tropidothorax elegans SNMP1 and SNMP2 with SNMPs from other insects
Note: the transmembrane domains are boxed; the six conserved cysteines are marked with an asterisk; Abbreviated species names: Tele, Tropidothorax elegans; Aips, Agrotis ipsilon; Alin, Adelphocoris lineolatus; Slit, Spodoptera litura; Btab, Bemisia tabaci. ![]()
图 2
红脊长蝽SNMPs与其他昆虫SNMPs氨基酸序列的系统进化树
Figure 2.
Phylogenetic tree of the SNMPs of Tropidothorax elegans and other insects based on amino acid sequences by using neighbor-joining method
![]()
图 3
红脊长蝽SNMPs在不同组织中的相对表达量
注:An, 触角; He, 头; Th, 胸; Ab, 腹; Le, 足; Wi, 翅; 柱上不同字母表示不同组织间基因表达存在显著差异(P < 0.05). Figure 3.
Expression level of SNMPs in different tissues of Tropidothorax elegans
Note:An, antennae; He, heads; Th, thoraxes; Ab, abdomen; Le, legs; Wi, wings; The different letters above bars indicate there is a significant difference between different tissues (P < 0.05). -
[12] Levy E, Spahis S, Sinnett D, et al. Intestinal cholesterol transport proteins: an update and beyond[J]. Current Opinion in Lipidology, 2007, 18(3): 310-318.
DOI: 10.1097/MOL.0b013e32813fa2e2[8] Yang K, Huang L Q, Ning C, et al. Two single-point mutations shift the ligand selectivity of a pheromone receptor between two closely related moth species[J]. eLife, 2017, 6: e29100.
DOI: 10.7554/eLife.29100[27] Herboso L, Talamillo A, Pérez C, et al. Expression of the scavenger receptor class B type I (SR-BI) family in Drosophila melanogaster[J]. International Journal of Developmental Biology, 2011, 55(6): 603-611.
DOI: 10.1387/ijdb.103254lh[16] Forstner M, Gohl T, Gondesen I, et al. Differential expression of SNMP-1 and SNMP-2 proteins in pheromone sensitive hairs of moths[J]. Chemical Senses, 2008, 33(3): 291-299.
DOI: 10.1093/chemse/bjm087[33] Shan S, Wang S N, Song X, et al. Molecular characterization and expression of sensory neuron membrane proteins in the parasitoid Microplitis mediator (Hymenoptera: Braconidae)[J]. Insect Science, 2020, 27: 425-439.
DOI: 10.1111/1744-7917.12667[19] Hu Y Y, Xu S F, Wubie A J, et al. Molecular characterization and tissue localization of sensory neuron membrane protein from Chinese honey bee, Apiscerana cerana (Hymenoptera: Apidae)[J]. Applied Entomology and Zoology, 2013, 48(4): 533-545.
DOI: 10.1007/s13355-013-0215-1[4] Benton R, Vannice K S, Gomez-Diaz C, et al. Variant ionotropic glutamate receptors as chemosensory receptors in Drosophila[J]. Cell, 2009, 136(1): 149-162.
DOI: 10.1016/j.cell.2008.12.001[5] Leal W S. Odorant reception in insects: roles of receptors, binding proteins, and degrading enzymes[J]. Annual Review of Entomology, 2013, 58: 373-391.
DOI: 10.1146/annurev-ento-120811-153635[24] Huang X L, Liu L, Fang Y Q, et al. Expression of a sensory neuron membrane protein SNMP2 in Olfactory Sensilla of codling moth Cydia pomonella (Lepidoptera: Tortricidae)[J]. Journal of Economic Entomology, 2016, 4(109): 1907-1913.
[30] 胡颖颖, 徐书法, 李 微, 等. 中华蜜蜂感觉神经元膜蛋白基因克隆、组织表达分析及原核表达[J]. 昆虫学报, 2013, 56(1):9-17.
[25] Sun L, Wang Q, Zhang Y, et al. Expression patterns and colocalization of two sensory neurone membrane proteins in Ectropis oblique Prout, a geometrid moth pest that uses Type-II sex pheromones[J]. Insect Molecular Biology, 2019, 28(3): 342-354.
DOI: 10.1111/imb.12555[3] Zhang J, Walker W B, Wang G. Pheromone reception in moths: from molecules to behaviors[J]. Progress in Molecular Biology and Translational Science, 2015, 130: 109-128.
[22] Song Y Q, Sun H Z, Du J. Identification and tissue distribution of chemosensory protein and odorant binding protein genes in Tropidothorax elegans Distant (Hemiptera: Lygaeidae)[J]. Scientific Reports, 2018, 8: 7803.
DOI: 10.1038/s41598-018-26137-6[13] Vogt R G, Miller N E, Litvack R, et al. The insect SNMP gene family[J]. Insect Biochemistry and Molecular Biology, 2009, 39(7): 448-456.
DOI: 10.1016/j.ibmb.2009.03.007[14] Martin C, Chevrot M, Poirier H, et al. CD36 as a lipid sensor[J]. Physiology & Behavior, 2011, 105: 36-42.
[32] Liu C, Zhang J, Liu Y, et al. Expression of SNMP1 and SNMP2 genes in antennal sensilla of Spodoptera exigua (Hübner)[J]. Archives of Insect Biochemistry and Physiology, 2014, 85(2): 114-126.
DOI: 10.1002/arch.21150[2] Sun L, Xiao H J, Gu S H, et al. Perception of potential sex pheromones and host-associated volatiles in the cotton plant bug, Adelphocoris fasciaticollis (Hemiptera: Miridae): morphology and electrophysiology[J]. Applied Entomology and Zoology, 2014, 49(1): 43-57.
DOI: 10.1007/s13355-013-0223-1[9] Rogers M E, Sun M, Lerner M R, et al. Snmp-1, a novel membrane protein of olfactory neurons of the silk moth Antheraea polyphemus with homology to the CD36 family of membrane proteins[J]. Journal of Biological Chemistry, 1997, 272(23): 14792-14799.
DOI: 10.1074/jbc.272.23.14792[6] Brito N F, Moreira M F, Melo A C A. A look inside odorant-binding proteins in insect chemoreception[J]. Journal of Insect Physiology, 2016, 95: 51-65.
DOI: 10.1016/j.jinsphys.2016.09.008[21] Zhang H J, Xu W, Chen Q M, et al. A phylogenomics approach to characterizing sensory neuron membrane proteins (SNMPs) in Lepidoptera[J]. Insect Biochemistry and Molecular Biology, 2020, 118: 103313.
DOI: 10.1016/j.ibmb.2020.103313[36] Liu S, Qiao F, Liang Q M, et al. Molecular characterization of two sensory neuron membrane proteins from Chilo suppressalis (Lepidoptera: Pyralidae)[J]. Annals of the Entomological Society of America, 2013, 106(3): 378-384.
DOI: 10.1603/AN12099[1] Kaupp U B. Olfactory signalling in vertebrates and insects: differences and commonalities[J]. Nature Reviews Neuroscience, 2010, 11(3): 188-200.
DOI: 10.1038/nrn2789[17] Jin X, Ha T S, Smith D P. SNMP is a signaling component required for pheromone sensitivity in Drosophila[J]. Proceedings of the National Academy of Sciences of the United States of America, 2008, 105(31): 10996-11001.
DOI: 10.1073/pnas.0803309105[7] de Fouchier A, Walker W B III, Montagné N, et al. Functional evolution of Lepidoptera olfactory receptors revealed by deorphanization of a moth repertoire[J]. Nature Communications, 2017, 8: 15709.
DOI: 10.1038/ncomms15709[11] Febbraio M, Silverstein R L. CD36: implications in cardiovascular disease[J]. The International Journal of Biochemistry & Cell Biology, 2007, 39(11): 2012-2030.
[38] Montell C. A taste of the Drosophila gustatory receptors[J]. Current Opinion in Neurobiology, 2009, 19(4): 345-353.
DOI: 10.1016/j.conb.2009.07.001[28] Pregitzer P, Greschista M, Breer H, et al. The sensory neuron membrane protein SNMP1 contributes to the sensitivity of a pheromone detection system[J]. Insect Molecular Biology, 2014, 23(6): 733-742.
DOI: 10.1111/imb.12119[35] Li P Y, Qin Y C. Molecular cloning and characterization of sensory neuron membrane protein and expression pattern analysis in the diamondback moth, Plutella xylostella (Lepidoptera: Plutellidae)[J]. Applied Entomology and Zoology, 2011, 46(497): 497-504.
[10] Rogers M E, Krieger J, Vogt R G. Antennal SNMPs (sensory neuron membrane proteins) of lepidoptera define a unique family of invertebrate CD36-like proteins[J]. Journal of Neurobiology, 2001, 49(1): 47-61.
DOI: 10.1002/neu.1065[20] Zhang H J, Xu W, Sun L N, et al. Functional characterization of sensory neuron membrane proteins (SNMPs)[J]. BioRxiv. 2018, Available at: https://doi. org/10.1101/262154.
[15] Rogers M E, Steinbrecht R A, Vogt R G. Expression of SNMP-1 in olfactory neurons and sensilla of male and female antennae of the silkmoth Antheraea polyphemus[J]. Cell and Tissue Research, 2001b, 303(3): 433-446.
DOI: 10.1007/s004410000305[37] Liu S, Zhang Y R, Zhou W W, et al. Identification and characterization of two sensory neuron membrane proteins from Cnaphalocrocis medinalis (Lepidoptera: Pyralidae)[J]. Archives of Insect Biochemistry and Physiology, 2013, 82(1): 29-42.
DOI: 10.1002/arch.21069[29] Gu S H, Yang R N, Guo M B, et al. Molecular identification and differential expression of sensory neuron memebrane proteins in the antennae of the black cutworm moth Agrotis ipsilon[J]. Journal of Insect Physiology, 2013, 59: 430-443.
DOI: 10.1016/j.jinsphys.2013.02.003[31] Leal W S, Ishida Y, Pelletier J, et al. Olfactory proteins mediating chemical communication in the navel orangeworm moth, Amyelois transitella[J]. PLoS One, 2009, 4(9): e7235.
DOI: 10.1371/journal.pone.0007235[26] Yang H Y, Ning S Y, Sun X, et al. Identification and characterization of two sensory neuron membrane proteins from Onion Maggot (Diptera: Anthomyiidae)[J]. Journal of Economic Entomology, 2020, 113(1): 418-426.
[23] Livak K J, Schmittgen T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-∆∆CT method[J]. Methods, 2001, 25(4): 402-408.
DOI: 10.1006/meth.2001.1262[18] Nichols Z, Vogt R G. The SNMP/CD36 gene family in Diptera, Hymenoptera and Coleoptera: Drosophila melanogaster, D. pseudoobscura, Anopheles gambiae, Aedes aegypti, Apis mellifera, and Tribolium castaneum[J]. Insect Biochemistry and Molecular Biology, 2008, 38(4): 398-415.
DOI: 10.1016/j.ibmb.2007.11.003[34] Benton R, Vannice K S, Vosshall L B. An essential role for a CD36-related receptor in pheromone detection in Drosophila[J]. Nature, 2007, 450(7167): 289-293.
DOI: 10.1038/nature06328
 下载:
下载:

计量
- 文章访问数: 3955
- HTML全文浏览量: 2860
- PDF下载量: 48
 下载:
下载:












